
Search Your Topic
Translation- an overview
The pathway of protein synthesis is called Translation because the ‘language’ of the nucleotide sequence on the mRNA is translated into the language of the amino acid sequence. The m RNA is translated from its 5’end to its 3’end, producing a protein synthesized from its amino-terminal end to its carboxyl-terminal end.
Prokaryotic Translation
Components required for Translation-
- Amino acids
- Transfer RNA
- Messenger RNA
- Aminoacyl t RNA synthetase
- Functionally competent ribosomes
- Protein factors
- ATP and GTP as a source of energy
Ribosomes
Ribosomes are large complexes of protein and r RNA. They consist of two subunits one large (Heavy) and one small (Light) whose relative sizes are generally given in terms of their sedimentation coefficients or S (Svedberg) values (figure-1). The S values are determined by the shape as well as by the molecular mass; their numerical values are strictly not additive. The prokaryotic 50S and 30S ribosomal subunits together form a ribosome with an S value of 70.

Figure-1- Ribosome is composed of two subunits one large (Heavy) and one small (Light);
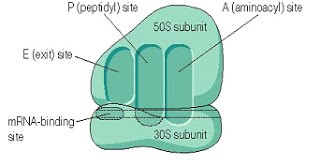
Figure-2-A and P sites of a Ribosome
The ribosome has two binding sites for t RNA molecule A and P sites, each of which extends over both subunits. Together they cover the neighboring codons. (See figure-2)
During translation, The A site binds an incoming Aminoacyl t RNA as directed by the codon currently occupying the site. This codon specifies the next amino acid to be added to the growing peptide chain.
The P site codon is occupied by the Peptidyl-t RNA. This t RNA carries the chain of amino acids that have already been synthesized.
An E site is also there that is occupied by the empty t RNA that is about to exit the ribosome
Transfer RNA
At least one specific type of t RNA is required per amino acid. Two sites are important, one Amino acid attachment site the other is Anticodon site. Because of their ability to carry a specific amino acid and to recognize the codons for that amino acid, t RNAs are called the adapter molecules.

Figure-3-Secondary structure of Transfer RNA
Amino acids are activated before incorporation and this activation is brought about by aminoacyl t RNA synthetase in the presence of ATP. There is at least one aminoacyl t RNA synthetase per amino acid. The carboxyl group of the amino acid is esterified to the 3’hydroxyl group of the t RNA.
Steps of Protein Synthesis
The process of protein synthesis is divided into 3 stages
i) Initiation
ii) Elongation
iii) Termination
i) Initiation-Initiation of the protein synthesis involves the assembly of the components of the translation system before the peptide bond formation occurs. These components include
o Two ribosomal subunits
o m RNA
o Aminoacyl tRNA specified by the codons in the message
o GTP and
o Initiation factors that facilitate the assembly of this initiation complex. In prokaryotes, three initiation factors are known
(IF-1,IF-2 and IF-3) while in eukaryotes there are at least 9 designated as e- IF to indicate the eukaryotic origin
Ribosomal assembly and formation of the initiation complex. The small ribosomal subunit binds to Initiation Factor 3 (IF3). The small subunit/IF3 complex binds to the mRNA. Specifically, it binds to the sequence AGG AGG, known as the Shine-Dalgarno sequence, which is found in all prokaryotic mRNAs. (Figure 4).

Figure-4- showing Shine Dalgarno sequence present in mRNA.
Meanwhile, the fmet tRNA binds to Initiation Factor 2 (IF2), which promotes binding of the tRNA to the start codon (figure 5).

Figure-5- showing the binding of f Met tRNA with IF-2
The small subunit/IF3 complex scans along the mRNA until it encounters the start codon. The tRNA/IF2 complex also binds to the start codon. This complex of the small ribosomal subunit, IF3, initiator tRNA, and IF2 is called the initiation complex. (Figure-6)

Figure-6- showing the formation of the initiation complex.

Figure-7- showing the formation of the initiation complex and release of IF2
At this point, the large ribosomal subunit joins in. A molecule of GTP is hydrolyzed, and the initiation factors are released. The ribosomal complex is now ready for protein synthesis.(Figure-7)
When the ribosome is assembled, two tRNA binding sites are created; these are designated ‘P’ and ‘A’ (P stands for Peptidyl, A stands for Aminoacyl). The initiator tRNA is in the P site, and the A site will be filled by the tRNA with the anticodon that is complementary to the codon next to the start. (In this case, it is the tRNA that binds proline.) Figure-8

Figure-8- showing the A and P sites
ii) Elongation
When the second tRNA base pairs with the appropriate codon in the mRNA, an enzyme called Peptidyl transferase catalyzes the formation of a peptide bond between the two amino acids present (while breaking the bond between fmet and its tRNA).This activity is intrinsic to the 23S r RNA found in the large subunit. Since the r RNA catalyzes this process, it is referred to as the Ribozyme. At this point, the whole ribosome shifts over one codon. This shift requires several elongation factors (not shown) and energy from the hydrolysis of GTP. The result of the shift is that the uncharged tRNA that was in the P site is ejected, and the tRNA that was in the A site is now in the P site. The A site is free to accept the tRNA molecule with the appropriate anticodon for the next codon in the mRNA.(Figure 9)

Figure-9- showing the process of elongation
The next tRNA base pairs with the next codon and Peptidyl transferase catalyze the formation of a peptide bond between the new amino acid and the growing peptide chain. Once again, the ribosome shifts over, so that the uncharged tRNA is expelled, and the tRNA with the peptide chain occupies the P site. (This is why this site is called the ‘Peptidyl’ site – after the shift, it contains the tRNA with the growing peptide chain. The other site will accept a tRNA with an amino acid, hence the name ‘Aminoacyl’ site.) The process of shifting and peptide bond formation continues over and over until a termination codon is encountered (figure-10).

Figure-10- showing the process of termination
The elongation process is fairly rapid, with prokaryotic ribosomes able to add 15 amino acids to the growing polypeptide every second. The process is also relatively error-free. Only one mistake is made every 10,000 amino acids. For large proteins of 1000 amino acids, that would mean one wrong amino acid in every 10 polypeptides.
iii) Termination
When a termination codon enters the A site, translation halts. This is because there is no tRNA with an anticodon that is complementary to any of the stop codons. The release factor causes the translation complex to fall apart and cleaves the polypeptide from the final tRNA (figure-11).

Figure-11- Showing the releasing factor occupying the stop codon.
The polypeptide product is now free to function in the cell. The mRNA molecule is now available to be translated again. Very often, more than one ribosome will translate a single mRNA at the same time. One ribosome will initiate translation, and after it moves down the mRNA a bit, another ribosome will initiate, then another, and so on. The structure consisting of multiple ribosomes translating a single mRNA molecule is called a polysome. Eventually, the mRNA is degraded, and translation of that particular message will cease.
Eukaryotic Translation
The eukaryotic translation is very similar overall to prokaryotic translation. There are a few notable differences, These include the following:
Eukaryotic mRNAs do not contain a Shine-Dalgarno sequence. Instead, ribosomal subunits recognize and bind to the 5′ cap of eukaryotic mRNAs. In other words, the 5′ cap takes the place of the Shine-Dalgarno sequence.
Eukaryotes do not use formyl methionine as the first amino acid in every polypeptide; ordinary methionine is used. Eukaryotes do have a specific initiator tRNA, however, Eukaryotic translation involves many more protein factors than prokaryotic translation (For example, eukaryotic initiation involves at least 10 factors, instead of the 3 in prokaryotes).
Inhibitors of protein synthesis
- The tetracyclines (tetracycline, doxycycline, demeclocycline, minocycline, etc.) block bacterial translation by binding reversibly to the 30S subunit and distorting it in such a way that the anticodon of the charged tRNAs cannot align properly with the codons of the mRNA.
- Puromycin structurally binds to the aminoacyl t RNA and becomes incorporated into the growing peptide chain thus causing inhibition of further elongation.
- Chloramphenicol inhibits prokaryotic Peptidyl Transferase
- Clindamycin and Erythromycin bind irreversibly to a site on the 50 s subunit of the bacterial ribosome thus inhibit translocation.
- Diphtheria toxin inactivates the eukaryotic elongation factors thus prevent translocation.
Post Translational Modifications
The newly synthesized protein is modified to become functionally active. The various post-translational modifications are as follows
1) Trimming- Trimming removes excess amino acids.
2) Covalent Modifications
a) Phosphorylation
b) Glycosylation
c) Hydroxylation
d) Gamma carboxylation
e) Isoprenylation
f) Methylation
g) Acetylation
h) Protein degradation
- Phosphorylation may activate or inactivate the protein.e.g.Glycogen Phosphorylase becomes active while glycogen synthase becomes inactive on phosphorylation.
- Glycosylation targets a protein to become a part of the plasma membrane, or lysosomes or be secreted out of the cell
- Hydroxylation such as seen in collagen is required for acquiring the three-dimensional structure and for imparting strength.
- Gamma carboxylation of glutamic acid residues of prothrombin takes place in the presence of vitamin K.
- Methylation or Acetylation of histones takes place for gene expression.
- Defective proteins or destined for turn over are marked for destruction by attachment of a Ubiquitin protein. Proteins marked in this way are degraded by a cellular component known as the Proteasome.
3) Subunit Aggregation- Examples are immunoglobulins, hemoglobin, and maturation of collagen. The failure of post-translational modifications affects the functional capacity of the proteins.